In the late 1990's student interns at the NDB developed a Music Atlas to provide a unique way to explore structures of DNA. The full Atlas is replicated here. Two types of musical representations were recorded. Project 1 focuses on B-DNA base pairing, while Project 2 uses a more detailed scheme to represent DNA conformation, mismatches, modifications, and more. Click on the tabs below for more information about Projects 1 and 2, and to listen to or download compositions arranged for A-DNA, B-DNA, Z-DNA, and mispair (A- or B-DNA) sequences. Only B-DNA compositions have WAV format files that can be listened to in-browser; other formats (au,mid) must be downloaded for listening in an application (see Listening Tips).
By Joanna de la Cruz (RU '98)
DNA can be represented in a variety of ways, which can provide different visual perspectives of molecular structure. DNA can also be aurally represented when a correspondence is created between the bases and musical notes, as seen in Dr. David Deamer's earlier work DNA Suite (1983). This Musical Atlas presents an aural representation of the B-DNA molecules without mismatches, drugs, or modifiers that were contained in the NDB as of July 25, 1996.
For each structure, there is a "Plain Melody," which follows a simple algorithm to highlight the structure's sequence, and a "Composition," which follows a more complicated algorithm that features the base pairing of the structure. These algorithms contain rules for determining the meter and rhythm for each piece. In each case, the notes are assigned to each base as follows:
| Adenine | A | Cytosine | C | Thymine | E | Guanine | G |
The notes are found within the range a to g1. Each base of the asymmetric strand is given one note. The strand is read 5' to 3'. Thymine was assigned to E so that the four bases would fall into an a minor seventh chord.
In each melody, each base in the sequence is played for one beat. If there are four adenines in a row, the note A will be played four (separate) times. If there is only one cytosine, it will be played for one beat, etc. In these melodies, the sequence for each structure is played once. The purpose of this Melody is to provide the listener with a 'feel' for the nucleotide without accompaniment.

Figure 1. Plain Melody for BDF062, Strand A (C G C T G G)
For each composition, there are four measures in which every quarter note gets one beat. The number of beats per measure is based upon the length of the nucleotide; the number of beats per measure is half the number of bases per strand. For example, if the nucleotide is 12 base pairs long, then the number of beats per measure will be 6 (time signature = 6/4). The sequences used here all had an even number of base pairs. However, if a sequence contained an odd number of bases, the number of beats per measure will be half that amount minus the remainder. For example, a 13 base pair piece would get 6 beats per measure.
Each base in the asymmetrical strand is an eighth note (as opposed to the quarter note used in the Plain Melody; an eighth note is half the length of a quarter note). The compositions consist of two lines:
| Base from asymmetrical strand | Note Played in Melodic Line (Strand Sequence) |
Note Played in Bass Line (Complementary Strand Sequence) | Assigned note to follow Base Pairing note in Bass Line | Range for base pairs and assigned bass note |
The base pair will play at the same time as the base in the asymmetrical strand but will only play for half the amount of time given in the second measure. It will then be followed by the new note assigned by the algorithm having the same time length as the base pair preceding it. For example, using the same sequence as above, C G C A A A T T T G C G, cytosine's note is a C. When the C plays for half a beat (eighth note) a G plays in the bass line for one quarter of a beat (sixteenth note) followed by an F which is also played for one quarter of a beat (sixteenth note). For the dotted quarter note A, the bass line will have an E playing for half that time followed by a D played for the same amount of time.
The fourth measure ends in a somewhat static chord progression. Each base in the melodic line is accompanied by its corresponding chord in the bass line. To create a stable chord, it was decided that these chords should be a fifth down from the base note, and should be in the same key signature (a minor).The chords assigned by the algorithm will play simultaneously with the corresponding bases in the top line. Chord inversions are acceptable as long as the line is kept as smooth as possible.

Composition for BDL038
Acknowledgements Thanks to Jason Young for providing technical assistance.
By Gautam Malhotra (RU '96)
"Seeing" things on the molecular level is obviously impossible with the naked eye, as even the electron microscope provides limited visibility of molecules. Therefore, other ways of "looking" at structures have been developed. Since the majority of us have difficulty visualizing molecular structure by glancing at tables of numbers, a familiar adage can be altered to "a picture is worth a lot of data." Thus drawings and models are a necessary tool for understanding and interpreting scientific data.
However, perceptions of reality are not limited only to vision but to four other senses. The concept of "hearing" DNA through music as seen in Dr. David Deamer's DNA Suite (1983), motivated Joanna de la Cruz (Rutgers University '98 of the NDB) to further develop aural representations of molecular structures to aid in "hearing" things which could not be "seen." De la Cruz created compositions based around DNA sequences to provide a unique representation of B-DNA sequence.
The algorithm presented here builds on Deamer's and de la Cruz's ideas in a number of ways. First, the new algorithm represents more aspects of the DNA structure (conformation, mismatches, modifications, nicks, uracil, inosine, etc). Secondly, it uses more than one timbre or tone color to aid in representation (i.e. the main sequence is represented by a piano while the complementary sequence is represented by a string section). The primary goal of this project was to use music to identify structures. Thus the representational value of each musical idea was more important than its aesthetic consequence.
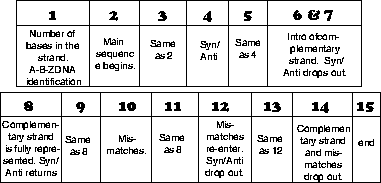
Measure 1: A number of equally spaced percussive clicks are sounded to represent the number of nitrogenous bases in the DNA strand. For example, if there are a total of 6 successive clicks, then there are 6 bases in the strand of DNA (and consequently the time signature is 6/8). The first sound establishes whether you are listening to A-, B-, or Z-DNA.
These sounds are repeated to signal the beginning of every new measure and were chosen purely for aesthetic reasons.
Measure 2: The melody, or main strand, is introduced by the piano and is repeated until the end of the piece. Notes are equidistant since the nitrogenous bases themselves are structurally an equal distance apart. Thus spaces in the melody can reveal missing bases and extra notes can represent hairpin loops and overhangs. Notational representation of bases is shown below.
A-DNA | B-DNA | Z-DNA | |
Adenine | A | a1 | a flat |
Cytosine | c | c2 | c1 |
Thymine | E | e1 | e flat |
Guanine | G | g1 | g |
Inosine | B | b1 | b flat |
Uracil | D | d1 | d |
Note: c1 is "middle c." c2 is one octave above "middle c" and c is one octave below it. Capitalized letters are in the octave directly below c.
A-DNA is represented by a low range of pitches which correspond to low anti conformation. B-DNA's high range represents high anti conformation. Z-DNA has pitches between A-DNA & B-DNA ranges, which represent sequence only. A-DNA & B-DNA are both right handed helices which are being represented in the major mode. Z-DNA is a left handed helix and is therefore represented by the natural minor mode. This explains why the notes A, B and E in Z-DNA have been flatted. (Note: the keys of C major and c minor were chosen for convenience and clarity of identification.) Thus the left/right handedness of a strand can be identified by listening to the mode of each piece.
Measure 3: Exact repetition of measure 2.
Measure 4: Syn/Anti configuration is represented by the choir voice.
Measure 5: Exact repetition of measure 4.
Measure 6 & 7: The strings introduce the complementary strand using the algorithm described below. This introduction develops for two measures. The complementary strand's melody will always sound an octave below that of the main strand. Syn/Anti choir voice drops out.
Measure 8: Full sequence of the complementary strand is established using the same principles for pitch and duration as were used for the main strand. The syn/Anti choir voice re-enters.
Measure 9: Exact repetition of measure 8.
Measure 10: If mismatches are present in the structure, they are represented in this measure by a layer of Orchestra Hits at the precise locations of the mismatches. (Orchestra Hits is a combination of strings, timpanis, brass, and woodwinds which achieves a loud staccato snap.) This measure will sound the same as measure 9 for structures without mismatches.
Measure 11: Exact repetition of measure 8.
Measure 12: If present in the structure, mismatches are represented again. The Syn/Anti choir voice drops out.
Measure 13: Exact repetition of measure 12.
Measure 14: The complementary strand and any mismatches drop out.
Measure 15: Muted triangle sound indicates the end of the piece. Anything sounded after this is structurally meaningless. (However, for the sake of simplicity these pieces consistently end with single note cadences.)
The complementary strand, which is represented by the String Section sound, is preceded by a 2 measure melodic introduction. This melody is the result of an algorithm which was used purely for aesthetic purposes to relieve some of the monotony of the music. It does not signify structure.

Acknowledgements Thanks to Joanna de la Cruz for helpful discussion.
The most important requirement to listening to these files is having hardware that can listen to .au, .wav, and .mid files. Then, the appropriate software is needed. Finally, your WWW browser should be set up so that the MIME type description is set to be handled by the appropriate player.
Unlike MIDI files, audio files are actual sound recordings. These recordings are offered in two formats (.au and .wav) to accommodate different systems. Except for the format type, these files are identical. To hear these files, be sure that your browser matches the appropriate software with the MIME type for .au and .wav files.
The MIME type of the .mid files on this Atlas is audio/midi. If you are having trouble playing these files (e.g. your screen is filled with nonsense text) then you need to do one of the following:
Go to the general preferences (under Options) and look at Helpers. Make sure that a suitable application is chosen for MIME type audio/midi. This will enable your browser to call on a MIDI player when listening to a .mid file.
Quicktime is an appropriate application for Macintosh or Windows. After downloading a player, make sure you have selected it for audio/midi in the General Preferences.
If you prefer listening to the files on an external sound module this can be done by downloading the files and saving them as text or source files with the proper extension (i.e. .mid or .midi files). The instruments have been assigned general midi settings (GM) and should correspond to similar instruments universally.